植物は“危険サイン”により寄生線虫を察知する
2025年7月31日
理化学研究所農業・食品産業技術総合研究機構
科学技術振興機構(JST)
理化学研究所(理研)環境資源科学研究センター 植物免疫研究グループの白須 賢 グループディレクター(環境資源科学研究センター 副センター長)、門田 康弘 専任研究員、飯野 絵里香 研修生(研究当時)、農業・食品産業技術総合研究機構(農研機構)植物防疫研究部門 基盤防除技術研究領域の植原 健人 研究領域長らの国際共同研究グループは、植物が植物寄生線虫[1]のような動物型の病原体を分子レベルで認識する仕組みを初めて明らかにしました。
本研究成果は、世界中で甚大な農業被害を引き起こしている植物寄生線虫に対する免疫メカニズムの理解を深めるとともに、広範な病原体に抵抗性を持つ作物の開発に向けた新たな基盤の確立に貢献すると期待されます。
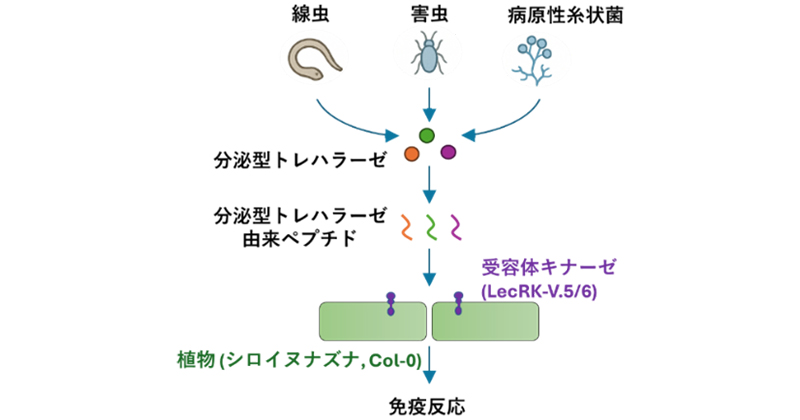
今回、国際共同研究グループは、植物寄生線虫が植物に寄生する際に分泌する酵素「トレハラーゼ[2]」の一部(ペプチド)を、植物が"危険サイン"として認識し、免疫反応を誘導する仕組みを解明しました。また、このサインを受け取るために植物が用いている受容体キナーゼ(リン酸化酵素)も同定しました。さらに、病原性糸状菌やアブラムシなどの害虫が持つトレハラーゼにも、同様のペプチドが存在し、植物がこれらにも同じ仕組みで反応することが分かりました。これらの成果から、植物が植物寄生線虫や病原性糸状菌、害虫など異なる病原体から放出される似た構造のペプチドを共通の危険サインとして認識し、防御反応を起こす仕組みが明らかになりました。
本研究は、科学雑誌『Science Advances』オンライン版(7月30日付)に掲載されました。
植物による分泌型トレハラーゼを介した植物寄生線虫、害虫、病原性糸状菌の認識機構
背景
植物が自然界で生き延びるためには、さまざまな病原体の侵入をいち早く察知し、それに応じて適切な防御反応を起こすことが不可欠です。植物は、病原体に共通して存在する特徴的な分子を目印として認識し、免疫反応を起動します。こうした目印は「微生物関連分子パターン(MAMPs)[3]」と呼ばれ、細菌のべん毛や真菌の細胞壁など、病原体に広く保存された構造の一部がこれに当たります。
植物の細胞表面には、MAMPsを検出するセンサーとして「パターン認識受容体(PRRs)[4]」が存在しており、これらがMAMPsを見つけると、免疫のスイッチが入り、防御反応が引き起こされます。これまでの研究により、病原性細菌や病原性糸状菌などの微生物に対するこの認識機構の多くが明らかにされてきました。
しかし、植物寄生線虫のような動物型の病原体に対して、植物がどのように反応し、それを見分けているのかは、長らく謎とされてきました。特に、植物寄生線虫由来のMAMPsに該当する分子や、それを感知するPRRsが存在するかどうかさえ不明であり、動物型病原体に対する植物免疫の理解は遅れていました。
研究手法と成果
植物寄生線虫を研究する上で大きな障壁となっていたのが、実験材料の安定的な確保でした。多くの植物寄生性の線虫は「絶対寄生性」と呼ばれ、増殖には植物を宿主として寄生することが不可欠です。そのため、生化学的な解析に必要な大量の植物寄生線虫を確保することが極めて困難であり、植物が植物寄生線虫をどのように認識しているのかを分子レベルで明らかにする研究は長らく進展していませんでした。
そこで国際共同研究グループは、寄生性ではない自由生活性線虫 Caenorhabditis elegans(C. elegans)を代替モデルとして活用しました。この線虫は実験室で大量培養が可能であり、植物への寄生能力がないにもかかわらず、C. elegansの抽出液はモデル植物シロイヌナズナに強い免疫反応を引き起こすことを発見しました。これを手掛かりに、抽出液から活性物質を精製した結果、「分泌型トレハラーゼ」と呼ばれる酵素の一部、わずか24個のアミノ酸から成るペプチド(TreCe24)が、強力な免疫反応誘導活性を持つことを明らかにしました(図1)。さらにこのペプチドは、根こぶ線虫(Meloidogyne incognita)をはじめ多様な植物寄生性の線虫のトレハラーゼにも高度に保存されており、植物がこれを危険のサインとして認識することで、植物寄生線虫の侵入をいち早く察知していることが分かりました。
図1 分泌型トレハラーゼ由来ペプチドによる免疫反応誘導
C. elegansの分泌型トレハラーゼに含まれる、トレハロース結合部位周辺由来のペプチドTreCe24は、シロイヌナズナCol-0において免疫反応を誘導する。この領域はさまざまな植物寄生線虫において高度に保存されている(A)。また、根こぶ線虫由来のペプチドTreMi31は免疫マーカー遺伝子であるCYP71A12を根の先端付近で強く誘導するとともに(B)、根に茶色物質の蓄積を誘導する(C)。この根の茶色化は、植物寄生線虫に対する免疫応答として他の植物種においても報告されている。さらにTreMi31は既知のMAMPsである細菌のべん毛タンパク質由来のペプチドflg22および糸状菌細胞壁由来のキチン(糸状菌に共通する細胞壁を構成する多糖類の一種)によって発現が誘導される遺伝子群のうち、半数以上の遺伝子の発現を誘導した(D)。これにより、TreMi31が植物の広範な免疫応答を引き起こす新規のMAMPとして機能する可能性が示唆された。
では、植物はこの共通ペプチドをどのように感知しているのでしょうか。
国際共同研究グループは、植物寄生線虫の中でも特に被害の大きい根こぶ線虫に由来し、TreCe24に類似するペプチド(TreMi31)を用いて、シロイヌナズナの複数の系統(アクセッション)[5]における反応の違いを調べました。その結果、TreMi31に全く反応しない非感受性系統Cvi-0を発見しました。この系統を手掛かりに遺伝学的解析を進めた結果、TreMi31に対する感受性を制御する遺伝子座が特定され、「LecRK-V.5」と呼ばれるレクチン受容体型キナーゼ遺伝子(LecRK)[6]が、ペプチド認識に必須であることが明らかになりました。感受性系統Col-0でLecRK-V.5を欠損した破壊株ではTreMi31による免疫応答(根の色素沈着や成長抑制など)が見られず、逆にLecRK-V.5を導入した相補株[7]では反応が回復したことから、この遺伝子が根こぶ線虫由来ペプチドの認識と免疫誘導に不可欠であることが確認されました(図2)。
図2 TreMi31により誘導される免疫反応に必要なLecRK-V.5
感受性系統Col-0のLecRK-V.5破壊株(lecrk-V.5-2)ではTreMi31による免疫マーカー遺伝子(CYP71A12)の発現が誘導されず(A)、根の茶色化(B)、および成長抑制も誘導されなかった(C)。一方、LecRK-V.5を導入した遺伝子相補株(lecrk-V.5-2/pLecRK-V.5:LecRK-V.5-3xHA)では、TreMi31によって根の茶色化(B)および成長抑制(C)が明確に誘導された。Bのスケールバー: 100マイクロメートル(μm、1μmは100万分の1メートル)。Cのスケールバー: 1cm。
さらに、LecRK-V.5と高い類似性を持つLecRK-V.6も、単独では弱いながらTreMi31に対する応答に関与し、LecRK-V.5を中心にLecRK-V.6が補助的に機能する、新たな免疫受容体群の存在が明らかになりました。
驚くべきことに、この分泌型トレハラーゼ由来ペプチドは、アブラムシなどの害虫や、Colletotrichum属の病原性糸状菌にも共通して存在することが判明しました(図3)。これらの生物に由来するペプチドを合成し植物に与えたところ、TreMi31と同様に免疫マーカー遺伝子の発現が強く誘導されました。その上、LecRK-V.5やLecRK-V.6を持たない植物ではこれらの反応が起こらないことから、植物が植物寄生線虫、害虫、病原性糸状菌といった異なるタイプの病原体に対して、共通の認識システムを用いて免疫応答を起こしている可能性が示されました。
このことは、シロイヌナズナが植物寄生線虫、害虫、病原性糸状菌といった異なるタイプの病原体に対して、共通の認識システムを利用して免疫応答を引き起こしている可能性を示唆しています。
図3 LecRK-Vs依存的に免疫を活性化する害虫、および病原糸状菌由来ペプチド
分泌型トレハラーゼ由来のペプチドは、病原性糸状菌および害虫においても高度に保存されている(A)。これらのペプチドは、シロイヌナズナCol-0において免疫マーカー遺伝子の発現を強く誘導したが、LecRK-V.5およびLecRK-V.6破壊株(lecrk-V.5/6/7-d)では誘導が認められなかった(B、C)。
今後の期待
本研究は、植物が植物寄生線虫のような動物型の病原体を分子レベルで認識する仕組みを初めて明らかにし、植物免疫の理解を大きく前進させる成果となりました。さらに、この認識機構が害虫や病原性糸状菌など他の病原体に対しても同様に働く可能性が示されたことは、植物が多様な敵に対して柔軟に対応するための基本的な免疫戦略の一端を示すものです。
LecRK-V.5やLecRK-V.6はアブラナ科の植物に特有の遺伝子ですが、これらを他の作物に導入することで、植物寄生線虫をはじめとする複数の病原体に対応できる新しいタイプの抵抗性作物の開発につながることが期待されます。
また、これまで植物の線虫に対する免疫反応は、寄生された部位に限られて起こるため、その仕組みを詳しく調べることが困難でした。今回発見したMAMPペプチドを使えば、植物全体で免疫反応を引き起こすことができ、線虫に対抗する防御物質を明らかにする手掛かりになると期待されます。
本研究成果は、国際連合が定めた「持続可能な開発目標(SDGs)[8]」のうち、「2.飢餓をゼロに」、「15.陸の豊かさも守ろう」に貢献するものです。
補足説明
- 1.植物寄生線虫植物の根に寄生して栄養を吸収する線形動物門に属する微小な動物である。体長はおおよそ1mm前後で、土壌中に生息し、植物の根に侵入して寄生生活を営む。イネ科・ナス科・マメ科など多様な作物を含む広範な宿主植物に寄生する。特に「根こぶ線虫(Meloidogyne属)」や「シストセンチュウ(Heterodera属、Globodera属)」は農業上の重要害虫であり、寄生により根の肥大や変形を引き起こし、養分吸収を阻害することで作物の生育を著しく低下させる。植物寄生線虫による世界の農業被害は、年間で推定800億ドル(約12兆円)以上に上るとされており、線虫対策は作物の安定生産において喫緊の課題となっている。
- 2.トレハラーゼ糖の一種であるトレハロースを加水分解し、グルコースに変換する酵素である。多くの微生物、線虫、昆虫などに保存されており、エネルギー供給やストレス耐性に関与する。中でも「分泌型トレハラーゼ」は、細胞外に分泌されるタイプのトレハラーゼであり、病原体が宿主環境中でトレハロースを利用する際に重要とされる。植物寄生線虫においては、シスト線虫(Heterodera属など)で分泌型トレハラーゼが病原性因子の候補として同定されており、寄生初期に植物の細胞外空間(アポプラスト)に放出されると考えられている。
- 3.微生物関連分子パターン(MAMPs)微生物に普遍的に存在し、宿主の免疫系によって認識される分子構造のこと。細菌のべん毛タンパク質、糸状菌の細胞壁分解産物であるキチン(糸状菌に共通する細胞壁を構成する多糖類の一種)などが含まれる。MAMPsはmicrobe-associated molecular patternsの略。
- 4.パターン認識受容体(PRRs)植物の細胞膜上に存在する免疫受容体であり、病原体に共通する分子パターン(MAMPs)を認識して免疫反応を誘導する役割を担う。病原菌由来の構造的特徴を直接検出し、初期の自然免疫を活性化する。代表的な例としては、細菌のべん毛タンパク質由来ペプチド(flg22)を認識するFLS2(Flagellin sensing 2)や、細菌の翻訳伸長因子EF-Tu由来ペプチド(elf18)を認識するEFR(EF-Tu receptor)が知られている。PRRsはpattern recognition receptorsの略。
- 5.シロイヌナズナの複数の系統(アクセッション)アクセッションとは、自然界から収集された個体群をもとに維持されている植物の遺伝的系統を指す。モデル植物であるシロイヌナズナ(Arabidopsis thaliana)では、世界中の異なる地域から採取された数百種以上のアクセッションが研究に用いられており、それぞれが異なる遺伝的背景や形質を示す。代表的なアクセッションとして、世界中で広く利用されている標準系統「Col-0(コロンビア)」がある。Col-0は遺伝子解析やトランスフォーメーション実験などに適しており、ゲノム解読の参照配列としても用いられている。一方、カーボベルデ諸島由来の「Cvi-0(カーボベルデアイランド)」は、Col-0とは異なる形態的・生理的特徴を持ち、遺伝的多様性の比較研究や遺伝子の機能解析に活用されている。本研究では、TreMi31に反応しない非感受性アクセッションとしてCvi-0を用いることで、ペプチド認識に関与する遺伝子(LecRK-V.5)の特定につながった。
- 6.レクチン受容体型キナーゼ遺伝子(LecRK)細胞外レクチン様ドメイン、膜貫通領域、細胞内キナーゼドメインから構成される受容体型タンパク質をコードし、植物の情報伝達に関与する。LecRKタンパク質は細胞外でリガンドを認識し、キナーゼ活性を介して細胞内にシグナルを伝える。
- 7.相補株特定の遺伝子が欠損または変異している植物に、その遺伝子を再導入することで元の機能を回復させた株のことである。この株を使う手法は、特定の遺伝子がある表現型の原因であるかどうかを検証するために用いられる。例えば、ある遺伝子の欠損変異体が特定の刺激に反応しない場合に、その遺伝子を再び導入して反応が回復すれば、その遺伝子がその反応に必要なことへの強力な証拠となる。
- 8.持続可能な開発目標(SDGs)2015年9月の国連サミットで採択された「持続可能な開発のための2030アジェンダ」にて記載された国際目標。持続可能な世界を実現するための17のゴールから構成され、地球上の誰一人として取り残さないことを誓っている。SDGsは発展途上国のみならず、先進国自身が取り組むユニバーサル(普遍的)なものであり、日本としても積極的に推進している(外務省ホームページから一部改変して転載)。
国際共同研究グループ
理化学研究所 環境資源科学研究センター 植物免疫研究グループ グループディレクター 白須 賢(シラス・ケン) (環境資源科学研究センター 副センター長) 専任研究員 門田 康弘(カドタ・ヤスヒロ) 研修生(研究当時)飯野 絵里香(イイノ・エリカ) テクニカルスタッフⅡ 槇 紀子(マキ・ノリコ) 基礎科学特別研究員(研究当時、現客員研究員)佐藤 一輝(サトウ・カズキ) 学振特別研究員PD 大野 恵梨佳(オオノ・エリカ) 研究員 石濱 伸明(イシハマ・ノブアキ) 基礎科学特別研究員 ブルーノ・ポクマン・ゴウ(Bruno Pok Man Ngou)
上級研究員 熊倉 直祐(クマクラ・ナオヨシ)
ケンブリッジ大学(英国)The Crop Science Centre 教授 セバスチャン・イーヴス・ファン・デン・アッカー(Sebastian Eves-van den Akker) 訪問学生 メイソン・ジェームズ・ルーゲン=ハンキー(Mason James Rugen-Hankey)
博士課程学生 スーユァン・ウェイ(Siyuan Wei)
Marc W Schmid社(スイス) 研究員 マーク・シュミッド(Marc W Schmid)
中部大学 応用生物学部 教授 鈴木 孝征(スズキ・タカマサ)
農研機構 植物防疫研究部門 基盤防除技術研究領域 研究領域長 植原 健人(ウエハラ・タケト)
研究支援
本研究は、日本学術振興会(JSPS)科学研究費助成事業基盤研究(S)「植物と病原体の攻防における分子機構(研究代表者:白須賢)」、同基盤研究(A)「アポプラストにおける植物-病原体相互作用(研究代表者:白須賢)」、同基盤研究(B)「植物による線虫認識機構の解明(研究代表者:門田康弘)」「統合ゲノミクスによる抗線虫活性を示す微生物群集のプロファイリングと農地での再構築(研究代表者:門田康弘)」「植物の膜貫通型受容体(様)キナーゼを介した植物寄生線虫の認識機構の解明(研究代表者:門田康弘)、同若手研究「エフェクターの網羅的解析による植物寄生線虫の感染戦略の解明(研究代表者:佐藤一輝)」「転写後制御のハイジャックによる植物寄生線虫の感染戦略の解明(研究代表者:佐藤一輝)」、同特別研究員奨励費「線虫に対する植物免疫機構の解明とその応用(研究代表者:飯野絵里香)」、同挑戦的研究(萌芽)「アグロバクテリウムのステルス化による植物の形質転換効率の向上(研究代表者:門田康弘)」、文部科学省科学研究費助成事業学術変革領域研究(A)「不均一環境変動に対する植物のレジリエンスを支える多層的情報統御の分子機構(研究領域代表者:松下智直)」の計画研究「不均一土壌環境に応答した寄生植物の感染統御機構(研究代表者:吉田聡子)」、科学技術振興機構(JST)革新的GX技術創出事業(GteX)「GXを駆動する微生物・植物「相互作用育種」の基盤構築(研究代表者:野村暢彦、JPMJGX23B2)」、同戦略的創造研究推進事業(ACT-X)「植物免疫誘導性の抗線虫物質とその生合成遺伝子の同定(研究代表者:佐藤一輝、JPMJAX2227)」、同先端国際共同研究推進事業(ASPIRE)「植物の誘導リプログラミングに立脚した新規バイオエコノミー基盤の創出(研究代表者:杉本慶子、JPMJAP2306)」、内閣府戦略的イノベーション創造プログラム(SIP)次世代農林水産業創造技術「持続可能な農業生産のための新たな総合的植物保護技術の開発(研究チームリーダー:植原健人、研究代表者:後藤千枝)」、RIKEN TRIPユースケースフィールドオミックスによる助成を受けて行われました。
原論文情報
- Erika Iino†, Yasuhiro Kadota†*, Noriko Maki, Erika Ono, Kazuki Sato, Nobuaki Ishihama, Bruno Pok Man Ngou, Marc W Schmid, Takamasa Suzuki, Taketo Uehara, Ken Shirasu* †These authors contributed equally to this work. *Corresponding authors, "A trehalase-derived MAMP triggers LecRK-V-mediated immune responses in Arabidopsis", Science Advances, 10.1126/sciadv.adv8896
発表者
理化学研究所 環境資源科学研究センター 植物免疫研究グループ グループディレクター 白須 賢(シラス・ケン) (環境資源科学研究センター 副センター長) 専任研究員 門田 康弘(カドタ・ヤスヒロ)
研修生(研究当時)飯野 絵里香(イイノ・エリカ)
農研機構 植物防疫研究部門 基盤防除技術研究領域 研究領域長 植原 健人(ウエハラ・タケト)
発表者のコメント
今回の研究では、免疫反応を引き起こす"鍵となるペプチド"と、それを受け取る植物側の受容体タンパク質の両方を特定することができました。異なる病原体に共通するペプチドを植物が一つの仕組みで見分けているという点に、植物の巧妙な防御戦略の一端を感じました。基礎研究として非常に面白いだけでなく、今後の応用展開にも期待が持てる成果だと感じています。(飯野 絵里香)
これまで謎の多かった植物による植物寄生線虫認識の仕組みの一端を明らかにすることができました。植物の植物寄生線虫に対する免疫反応は、寄生した部位でのみ局所的に誘導されるため、分子レベルでの解析が難しいという課題がありました。今回同定したMAMPペプチドと受容体キナーゼを用いることで、植物全体で免疫反応を誘導することが可能となり、植物寄生線虫に対する植物免疫の全体像を解明するための新たな道が開けたと考えています。(門田 康弘)
植物が、植物寄生線虫・病原性糸状菌・害虫といったまったく異なる生物群から共通の"危険サイン"を見分けているという点に、生物同士の進化的な駆け引きの結果としての巧妙な仕組みが感じられます。植物寄生線虫を含む異なる病原体に対して、同じ認識機構が用いられている可能性が示されたことで、新たな視点から抵抗性作物の開発を考える手掛かりが得られたと感じています。(植原 健人)
これまで病原性細菌や病原性糸状菌に対する植物の免疫応答は多くの知見が蓄積されてきましたが、動物に分類される植物寄生線虫に対して、植物がどのように反応しているのかは長らく不明でした。今回、線虫由来のペプチドを"危険のサイン"として感知する植物のセンサー(免疫受容体)を特定できたことで、動物型病原体に対する植物免疫研究の新たな道が開けたと考えています。(白須 賢)
JST事業に関する問い合わせ
科学技術振興機構 未来創造研究開発推進部 GteX推進グループ 波羅 仁(はら・まさし) Tel: 03-3512-3543
Email: [email protected]
報道担当
理化学研究所 広報部 報道担当 お問い合わせフォーム
農研機構 お問い合わせ | 農研機構
科学技術振興機構 広報課 Tel: 03-5214-8404
Email: [email protected]
産業利用に関するお問い合わせ
関連記事:



